Epigenetics of T lymphocytes in health and disease
DOI: https://doi.org/10.4414/smw.2015.14191
Cristina
Leoni, Lucia
Vincenzetti, Stefan
Emming, Silvia
Monticelli
Summary
The risk of developing autoimmune diseases depends on both genetic and environmental factors, with epigenetic mechanisms of regulation potentially translating environmental cues into stable modifications in gene expression. Such stable memory of a functional state has been deciphered into a number of molecular mechanisms that collectively define the epigenetic status of a cell. In recent years, it has become increasingly clear that epigenetic modifications are highly dynamic and are able to adapt to the changing environment, with important impact on the onset and development of a number of diseases. Here, we describe some of the epigenetic mechanisms of regulation of cellular functional states in T lymphocytes, with a particular focus on DNA methylation. We will also discuss current knowledge on the role of epigenetics in autoimmunity and consider open questions in the field.
DNA methylation and hydroxymethylation dynamics in the regulation of gene expression
“Epigenetic” classically described inheritable phenotypes that are not the result of alterations in the DNA sequence [1, 2]. Here, however, we will utilise a broader interpretation of such a definition to include all mechanisms that can provide stability to a given phenotype, including those that can be important to impart short-term memory of an environmental signal [3, 4]. Mechanisms providing such cellular memory of gene expression include histone modifications, as well as regulation mediated by microRNAs (miRNAs) [3], although we will mainly focus on DNA methylation dynamics and their role in regulating functions of T lymphocytes during normal immune responses, as well as in autoimmunity.

Figure 1
DNA methylation and hydroxymethylation.Cytosine modifications in DNA have functional consequences on gene expression. (a)Cytosine can be methylated by DNMT enzymes to give rise to 5mC, which can be oxidised by TET enzymes to 5hmC and further products (5fC and 5caC). In an active demethylation pathway, these products of TET enzymatic activity are likely to be excised and repaired by the thymine-DNA glycosylase enzyme and base excision repair mechanisms to regenerate the unmodified cytosine. Methylated DNA can be recognised by MBD proteins that can recruit histone modifying complexes (HMTs and HDACs). (b) DNA methylation occurs at regulatory regions (such as promoters), but also in gene bodies. DNMT enzymes are responsible for 5mC deposition, often resulting in the transcriptional repression of the methylated promoter. TET proteins activity can lead to demethylation with a potential for transcriptional activation.
5caC = 5-carboxylcytosine; 5fC = 5-formylcytosine; 5hmC = 5-hydroxymethylcytosine; 5mC = 5-methylcytosine; BER = base excision repair; DNMT = DNA methyltransferase; HDAC = histone deacetylase; HMT = histone methyltransferase; MBD = methyl-CpG-binding domain protein; TDG = thymine-DNA glycosylase; TET = ten-eleven translocation
DNA methylation
DNA methylation occurs most commonly at the 5’ position of the cytosine ring in the context of CpG dinucleotides, and is mediated by DNA methyltransferase enzymes (DNMT1, DNMT3a, DNMT3b), which show variable affinities for unmethylated and hemimethylated DNA. The importance of proper DNA methylation throughout mammalian development is highlighted by the consequences of germline deletions of DNMTs: Dnmt1 and Dnmt3bdeletions in mice are embryonically lethal, and Dnmt3a-null mice die perinatally [5, 6].
The relevance of these enzymes can also be inferred from the fact that they are often found mutated in a variety of diseases. For example, DNMT3a is frequently mutated in a number of haematological malignancies, and upon transplantation of haematopoietic stem cells lacking DNMT3a, mice develop a spectrum of malignancies similar to those seen in patients with mutations in this enzyme, supporting the crucial role of DNA methylation in disease [7, 8]. Mutations in the gene encoding for DNMT3b are instead found in patients with ICF (immunodeficiency, centromere instability and facial anomalies), a recessive autosomal syndrome, and were shown to lead to reduced DNA methyltransferase activity [9].
Methylated DNA provides a platform for the binding of several methyl-CpG-binding domain (MBD) proteins, which can coordinate downstream processes by recruiting multiprotein complexes containing histone deacetylases (HDACs) and histone methyltransferases (HMTs), leading to chromatin modifications and alterations in gene expression [5, 10]. However, histone tails can also influence recruitment of DNMTs, which was in fact shown to be independent of pre-existing DNA methylation [11]. Indeed, methylation at lysine 4 of histone H3 (H3K4) was shown to inhibit DNMT3a recruitment [12, 13], and H3K36me3 modulated DNMT3b binding and consequent DNA methylation specifically to transcribed gene bodies [11]. Importantly, accessibility of selected transcription factors is also influenced by DNA methylation, since the methylation status of a binding site can define binding affinity and even specificity for a given transcription factor [14–17].
Despite such prominent progress in the field and decades of intensive studies, the precise role of DNA methylation in regulating gene expression is still not completely understood, and many concepts about DNA methylation have had to be reconsidered in recent years. DNA methylation was classically associated with gene silencing, and this is indeed usually the case at transposable elements and gene promoters, but methylation downstream of the transcriptional start site (TSS) usually does not block elongation [18]. Moreover, DNA methylation was not necessarily considered of general importance, since model organisms such as Drosophila melanogaster and Caenorhabditis elegans show very low or essentially no cytosine methylation [19]. However, while cytosine methylation represents the dominant DNA modification in humans, very recent work showed how organisms with virtually no cytosine methylation show significant levels of other DNA modifications, such as N6-methyladenine (6mA), further highlighting a possible widespread regulatory role of DNA methylation (reviewed in [20]). Finally, DNA methylation was thought to be substantially irreversible, other than through passive mechanisms of dilution, but the recent discovery of pathways leading to DNA demethylation have led to a revision of this idea.
An effort to describe the methylome of up to 17 eukaryotic genomes was able to show that DNA methylation is highly conserved in eukaryotes, and that is was likely present in a common ancestor [19, 21, 22]. Interestingly, methylation of the gene body was a highly conserved feature across genomes, with exons being usually more methylated than introns and with reduced methylation at both the promoter region and the 3’ end of genes. While gene expression was mostly inversely correlated with promoter methylation, gene body methylation showed a parabolic correlation with expression: modestly expressed genes are more likely to be methylated, while both high and low expressers are the least likely to be methylated [19, 21, 22]. This generally conserved pattern indicates a role of DNA methylation in regulating multiple steps of transcription, including initiation, elongation, termination and even splicing [19, 23]. Indeed, genome-wide studies showed that removal of one DNMT enzyme leads to both hypo- and hypermethylation at specific loci, underlying the complexity of the regulation mediated by DNA-modifying enzymes and further highlighting the fact that DNA methylation does not always equate to gene silencing [24, 25].
Overall, these studies clearly challenged the general dogma that DNA methylation invariably corresponds to transcriptional silencing, and put forward the idea that the final outcome on transcriptional regulation relates to the context and spatial distribution of DNA methylation [10]. Therefore, DNA methylation appears to be intimately associated with gene transcription.
Reversal of DNA methylation by hydroxylation
Once thought to be a stable chromatin modification, DNA methylation is now understood to have a much more dynamic nature, as its distribution is influenced by DNA demethylation processes, which can occur either passively, by dilution upon DNA replication at each cell division, or through an active process initiated by ten-eleven translocation (TET1-3) enzymes [26] (fig. 1). These enzymes are responsible for the catalytic conversion of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC) and to further oxidation products, including 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC) [27, 28]. Similarly to DNMT enzymes, TET proteins are frequently found mutated in a number of diseases. Specifically, mutations in the TET2gene are common in many haematological disorders, and mice lacking TET2 show expansion of the pool of haematopoietic stem cells, with a bias towards myelopoiesis, which in some cases can turn to myeloproliferative disorder [29–33].
Although the biological significance of such oxidation derivatives of 5mC remains to be fully elucidated, several lines of evidence have determined their importance in transcriptional regulation. Indeed, they are likely to be intermediates in processes of DNA demethylation and they modify the binding patterns of several chromatin regulators and transcription factors, thereby influencing transcription. A recent work undertook the challenge of investigating the identity of a large number of 5mC and 5hmC-interacting proteins in the mouse, revealing that each cytosine modification is able to recruit a distinct set of proteins with little overlap, and also that a large number of proteins preferentially interact with unmethylated cytosines [15]. These findings further indicate that the role of DNA methylation is integral to regulation (both positive and negative) of transcription. In general, because many of the methods used to study DNA methylation in the past could not distinguish between 5mC and 5hmC [34], it will be interesting to see how many of the functions attributed in the past to 5mC are actually dependent (or not) on the presence of 5hmC in the genome.
Epigenetic regulation of T cell functions
T lymphocytes are central to the orchestration of immune responses to invading pathogens, but their activity has to be carefully balanced and controlled to avoid tissue damage and pathology. Following antigenic stimulation, naïve T cells proliferate and differentiate into a number of effector and memory subsets, each characterised by the ability to produce specific signature cytokines. For example, CD4+ T helper (Th) cells differentiation to the Th1 subset is characterised by the production of high levels of interferon-gamma (IFNγ). Th2 cells instead mainly produce interleukin (IL)-4 and IL-13, while Th17 cells are characterised by the expression of IL-17. Th1 and Th17 cells in particular, and their associated cytokines, have been implicated in the pathogenesis of autoimmune diseases such as multiple sclerosis (MS) [35–37].
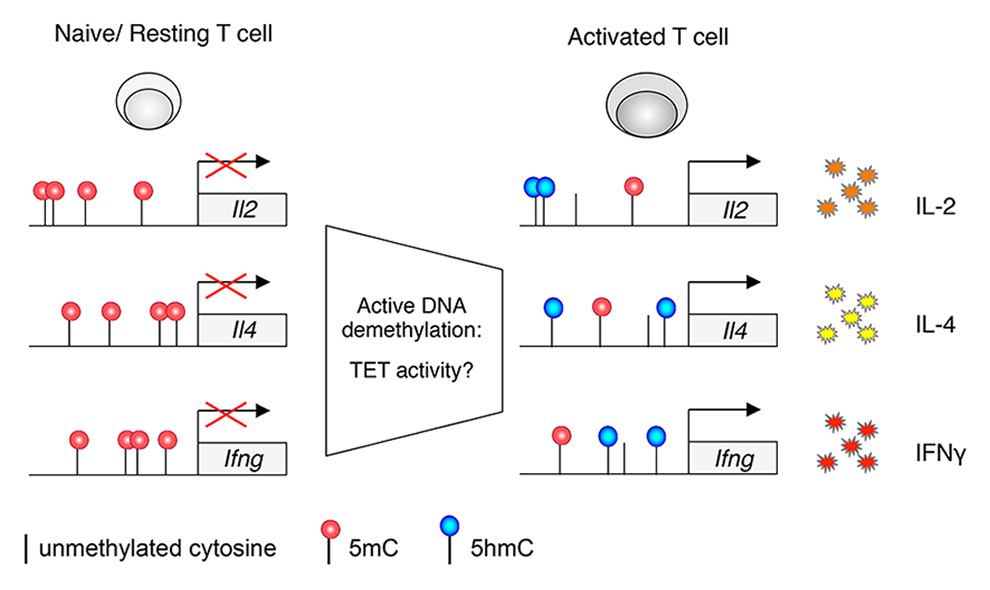
Figure 2
DNA methylation dynamics regulate cytokine production in T lymphocytes.DNA methylation modulates cytokine production upon T cell activation, and active demethylation processes (independent of cell replication) have been shown to act at some cytokine loci. Hypothetically, these processes may involve TET proteins as an underlying mechanism of demethylation.
5hmC = 5-hydroxymethylcytosine; 5mC = 5.methylcytosine; IFNγ = interferon-gamma; IL = interleukin; TET = ten-eleven translocation
Several genes controlling immune functions are known to be regulated by DNA methylation, although our understanding of the impact of epigenetic modifications in the regulation of T cell functions is currently still limited. The fact that DNA methylation can be dynamically modulated during T cell responses to an antigen is suggested by a number of observations. For instance, the Il2, Il4 and Ifng cytokine genes are known to be regulated by DNA methylation in T cells [38–40]. In the absence of DNMT1 or DNMT3a (but not DNMT3b), murine CD4+ T lymphocytes were unable to silence properly the Ifng and Il4loci under appropriate culture conditions, resulting in increased and promiscuous cytokine expression [41–43]; even T cells lacking MBD2 (methyl-CpG-binding domain protein 2), a reader of methylated DNA, showed a similar phenotype [44]. T cell activation in Th2-polarising conditions was shown to reduce substantially recruitment of DNMT1 to the Il4-Il13 locus in proliferating cells, eventually leading to reduced DNA methylation of this locus and enhanced gene expression [39]. Dysregulated Th2-type cytokine expression in the absence of DNMT1 was observed even in CD8+ T cells, indicating defective gene silencing [45]. Moreover, DNMT3a expression was shown in the mouse to be increased by T cell receptor (TCR) engagement. Accordingly, Dnmt3a-null murine T cells showed hypomethylated cytokine loci and dysregulated cytokine expression [41]. More recently, genome-wide methylation studies investigated changes in DNA methylation in effector and memory murine T cells compared with their naïve counterparts, highlighting the extent of chromatin remodelling events accompanying the acquisition of the effector/memory phenotype upon stimulation of a naïve T cell [46, 47].
Prior to the discovery of TET proteins, models for DNA demethylation placed emphasis on a passive process due to a failure of DNMT1 to methylate the newly synthesised DNA strand during the S phase of the cell cycle. However, the rapid and DNA replication-independent demethylation of the Il2 locus that was observed upon activation of CD4+ T cells and that causally affected Il2 gene transcription was inconsistent with such a model [40]. Along the same line, dynamic methylation of the Ifng locus was described for memory CD8+ T lymphocytes: while this locus was partially methylated at resting state, it underwent demethylation within 5 hours of antigenic stimulation [38]. This process was independent of DNA replication and cell division, and the involvement of a yet unidentified demethylase activity was proposed [38] (fig. 2). Interestingly, characterisation of the genome-wide distribution of 5hmC in mouse CD4+ T cells could not lead to a definitive answer about the role of 5hmC in T cell differentiation, although it showed a clear effect of TET2 in modulating effector responses in some T cell subsets, with reduced IL-10, IFNγ and IL-17A production in the absence of TET2 [48].
Among the various T cell subsets required to regulate immune responses properly, regulatory T cells (Tregs) are essential to suppress fatal autoimmunity throughout the lifetime of an organism. These cells are characterised by the expression of the transcription factor Foxp3, which plays a key role in their differentiation and function. Indeed, mutations or deficiency of the Foxp3gene lead to the development of autoimmune disease, causing the scurfy phenotype in mice and IPEX (immune dysfunction, polyendocrinopathy, X-linked) syndrome in humans [49]. Stable Foxp3 expression is essential to maintain Treg cell identity and functional integrity, and mechanisms are in place to preserve Treg cell lineage stability [50]. In particular, regulatory regions in the Foxp3 locus have to be maintained free of methylation in order to confer phenotypic stability to Tregs and, accordingly, inhibition of DNA methylation using 5-azacytidine or 5-aza-deoxycytidine led to stable Foxp3 expression [49–51]. By modulating the stability of the Treg phenotype, DNA methylation could therefore also have an impact on the onset of autoimmune diseases, and DNMT inhibitors were proposed as potentially useful tools to enhance Foxp3 expression in an attempt to limit inflammation [49]. Expression of DNMT1, but not DNMT3a, was, however, shown to be essential for the appropriate differentiation and function of Tregs, and mice with conditional deletion of DNMT1 in Tregs died of lethal autoimmunity [52]. DNA methylation therefore appears to have a complex role in regulating Treg stability and function that may hinder the development of epigenetic-based therapies.
DNA methylation in autoimmune and autoinflammatory diseases
Although the aetiology of MS and other autoimmune diseases is still largely unknown, it involves T cell-mediated processes [35]. Genome-wide association studies (GWAS) on MS patients have identified several genetic loci associated with a greater risk of developing the disease. Of these, the most commonly identified risk locus in MS and other autoimmune diseases is the major histocompatibility complex (MHC), pointing towards a fundamental role for antigen presentation, but many of the other loci associated with MS risk are related to immune functions, including cytokines and cytokine receptors, costimulatory receptors and signal transduction molecules [53].
Epigenetic mechanisms of regulation of gene expression may partly explain why only a proportion of genetically susceptible individuals may end up manifesting the disease. The complexity of such regulation and its impact on disease is exemplified by investigations on monozygotic twins: despite sharing identical genetic material, the risk of developing MS in a twin of an MS patient is only 25%, although it remains higher than in dizygotic twins or non-twin siblings [54]. Similarly, besides genetic predisposition, epigenetic factors appear to be important for the onset of systemic lupus erythematosus (SLE), since monozygotic twins are often discordant also for this disease [55]. To assess the relative contribution of heritable vs nonheritable influences on a variety of immunological parameters, a recent work investigated responses of 210 healthy twin pairs [56]. Interestingly, immune variation in this system was dominated by nonheritable determinants, some of which also became more divergent with age, suggesting a cumulative effect of environmental exposure [56].
Despite the clear evidence of an environmental component in disease development, it has not been easy to pinpoint the exact impact of epigenetics in autoimmune diseases; for example, a study investigating three monozygotic twin pairs discordant for MS found no reproducible differences in DNA methylation, single nucleotide polymorphisms or gene expression in CD4+ T cells [57]. A more recent study evaluated genetic and epigenetic determinants of 21 autoimmune diseases, observing that most causal variants map to enhancers rather than protein-coding genes and frequently coincide with regions bound by transcription factors and depleted of nucleosomes [58]. On the other hand, DNA methylation in cells of monozygotic twins discordant for the disease was also analysed in the case of type 1 diabetes (T1D), and the differentially methylated regions included several genes involved in immune responses [59]. Interestingly, some of the DNA methylation sites associated with T1D could be detected years before clinically overt T1D, suggesting a role in increasing the risk of developing this disease [59]. Changes in DNA methylation have been demonstrated also in SLE patients, with T lymphocytes from these patients showing overall genomic hypomethylation compared with controls [55]. Most importantly, exposing T cells to demethylating drugs resulted in the induction of a lupus-like disease in mice [60]. Although the mechanisms by which hypomethylated T cells induce SLE are not fully understood, recent evidence points towards a possible dominant role for IL-10 and IL-17 production in T cells, dependent on altered methylation status at these loci [61, 62]. Altered global DNA methylation and dysregulation at specific genetic loci have also been reported for rheumatoid arthritis (reviewed in [63, 64]), although a comprehensive view of the role of DNA methylation in autoimmunity and whether in each case it represents a cause or consequence is still lacking.
DNA methylation in disease: cause, consequence or correlation?
Aberrant DNA methylation is a pervasive feature of human tumours, which often show extensive genomic hypomethylation together with hypermethylation of tumour-suppressor genes (reviewed in [65]). As cancer progresses, the level of global methylation decreases, and DNA hypomethylation may contribute to cancer progression through chromosomal instability, reactivation of transposable elements and loss of imprinting [65]. Vice versa, hypermethylation of tumour-suppressor genes can have a major impact in suppressing the expression of these genes and therefore unleash uncontrolled cell proliferation. Similarly, global loss of 5hmC is a hallmark of a number of different tumours [66-68], although in spite of the diagnostic and prognostic implications, the clear biological significance of such altered methylation patterns remains to be fully understood. Importantly, alterations in the distribution of histone modifications have been described for a number of autoimmune diseases [63], and given the interplay between post-translational modification of histones and DNMT enzymes [69], at least part of the altered DNA methylation patterns observed in disease may potentially be linked to histone modifications.
While a causal link between mutations in DNA-modifying enzymes, altered methylation patterns and disease has been worked out in some cases [7, 8, 66], it has to be highlighted that some biological consequences of altered methylation do not necessarily link directly to disease. Indeed, changes in DNA methylation and hydroxymethylation are imperfectly maintained during replication and are subject to considerable epigenetic drift, that is, gradual increase or decrease at specific loci [70]. Such stochastic drift not only creates epigenetic mosaicism within populations, but it is also part of the normal process of aging, with aging cells and tissues showing considerable DNA hyper- or hypomethylation over time [70, 71]. Moreover, while some mouse models lacking DNMT3a or TET2 develop haematopoietic malignancies [7, 8, 30, 31, 33], this is not the case for mice lacking other DNMT or TET family members, suggesting that altered methylation patterns do not necessarily lead straight to disease. They can however predispose to its onset, and the lack of DNMT3a in particular appeared to confer flexibility to the methylome of haematopoietic stem cells, allowing cellular transformation after the acquisition of additional mutations [7, 8].
Interestingly, although the underlying mechanism remains unclear, chronic inflammation can be an important inducer of epigenetic drift, and potentially predisposing to the onset of disease. For example, Helicobacter pylori is a well-known inducer of chronic inflammation and gastric cancer, and altered patterns of DNA methylation are associated with H. pyloriinfection. In a gerbil model of gastric cancer, H. pyloriinfection was sufficient to induce DNA methylation in gastric epithelial cells, which could be partially reversed by H. pylorieradication or blocked by immunosuppressive drugs [72], suggesting that inflammation induced by the infection was a critical determinant of hypermethylation. Other determinants of methylation variations remain poorly understood [70].
|
Table 1: Common miRNAs dysregulated in autoimmune diseases. Examples of some common miRNAs that have been reported to influence autoimmunity in general, MS or its experimental model (EAE) in humans (Hsa), mouse (Mmu) or rat (Rno). |
|
miRNA
|
Expression change
|
Target/ Function
|
Model system
|
Outcome
|
PMID
|
| –
(Dicer/Drosha deletions) |
All miRNAs down |
Elimination of components of the miRNA machinery in Tregs |
Mmu |
Fatal autoimmunity |
18725525
18725526
18725527 |
| Let-7e |
Up |
Dysregulated cytokine production |
Mmu |
Silencing ameliorated EAE |
23079871 |
| miR-10 |
Up |
Bcl-6, T cell plasticity |
Mmu |
Delayed onset of EAE |
22544395 |
| miR-17~92 |
Up |
Treg functions |
Mmu |
Treg-specific loss of miR-17~92 results in exacerbated EAE |
23858035
20148420 |
| miR-20b |
Down |
Disease severity, Th17 responses |
Mmu |
Overexpression leads to reduced severity of EAE |
24842756 |
| miR-23b |
Down |
Suppresses inflammatory cytokine-mediated signaling |
Hsa/ Mmu |
Overexpression leads to reduced severity of EAE |
22660635 |
| miR-29 |
Up in MS and EAE |
T-bet, IFNg, Th1 differentiation |
Hsa/ Mmu |
Knock-out mice have reduced EAE |
22772450 |
| miR-132/212 |
Up |
Th17 differentiation |
Mmu |
Exacerbated EAE |
23818645 |
| miR-155 |
Up |
Positive regulator of inflammation |
Mmu |
Knock-out mice have reduced EAE |
20888269
21788439 |
| miR-181 |
Up |
Regulates threshold of TCR activation |
Hsa/ Rno |
Upregulated in CD4+ T cells of strains susceptible to EAE and in cerebrospinal fluid of MS patients |
17382377
25775432
23514736
23077021 |
| miR-301 |
Up |
Th17 differentiation |
Mmu |
Exacerbated EAE |
22517757 |
| miR-326 |
Up |
Disease severity, Th17 responses |
Hsa/ Mmu |
Expression correlates with disease |
19838199 |
MicroRNAs, immune regulation and autoimmunity
MicroRNAs have recently emerged as important regulators of both transcriptional and epigenetic programmes, with pivotal roles in regulating early T cell development, lineage specification and function [73]. As a result of their stability, miRNAs are most suitable to maintain memory of altered cellular states [3] and it is becoming increasingly clear that they are involved in regulating various aspects of autoimmunity (reviewed in [74, 75]), as well as in the regulation of DNA-modifying enzymes. For instance, miR-21 and miR-148a were shown to be highly expressed in patients with SLE, and to promote hypomethylation by repressing DNMT1 expression [76]. This in turn led to increased expression of autoimmune-associated genes like CD70 and LFA-1, surface markers that were shown to play a role in lupus [55].
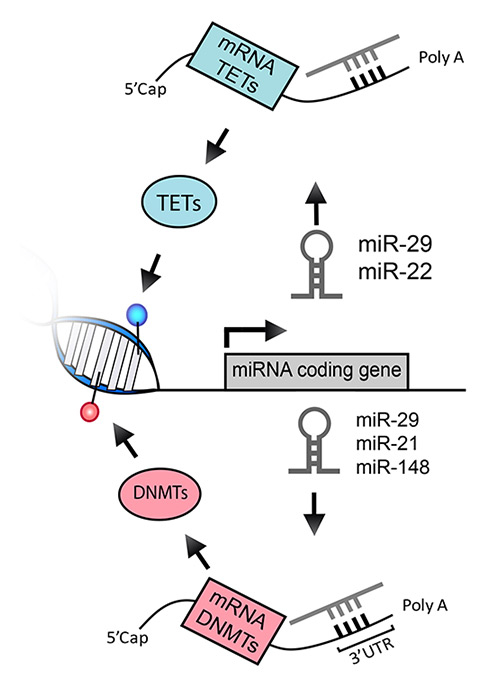
Figure 3
Interplay between miRNAs and the DNA methylation machineries. MiRNAs are both target and effector of epigenetic modifications. Expression of miRNAs can be modulated by levels of DNA methylation in the gene locus of any given miRNA. In turn, mature miRNAs bind the 3’UTR of target mRNAs leading to a reduction in protein output. DNMT and TET mRNAs can be targeted by miRNAs such as miR-29, miR-22 and others, thereby modulating the levels of DNA methylation and hydroxymethylation.
DNMT = DNA methyltransferase; miRNA = micro RNA; mRNA = messenger RNA; TET = ten-eleven translocation protein; UTR = untranslated region
A growing number of studies also identified miRNAs as potential novel therapeutic targets in animal models of MS as well as in primary cells derived from MS patients (table 1). For example, miR-155 and miR-29ab expression was found to be elevated in mouse CD4+ T cells during experimental autoimmune encephalomyelitis (EAE), and miR-155– or miR-29ab–deficient mice had a delayed course and reduced severity of disease [77, 78]. Another relevant example is provided by miR-326, whose expression was highly correlated with disease severity in patients with MS and in mice with EAE. Accordingly, modulation of miR-326 expression altered disease severity [79]. Vice versa, miR-20 was shown to be downregulated during EAE, and its overexpression led to reduced disease severity in mice [80].
In MS patients, distinct miRNA expression profiles were found in CD4+ and CD8+ T cells as well as B cells as compared with healthy donors, and miR-17-5p, another miRNA known to be involved in autoimmunity, was upregulated in CD4+ T cells from MS patients [81]. Further highlighting the importance of miRNAs in regulating various aspects of human disease, a role has even been proposed in the regulation of blood-brain barrier endothelial tightness in MS patients [82]. Moreover, miRNAs have been investigated as promising biomarkers for disease, being able to discriminate MS from other neurological diseases, as well as different disease courses of MS [83].
Despite encouraging results underscoring the role of miRNAs in autoimmune diseases, the biological function of most of these miRNAs and their relevance for disease in the context of MS is mostly unknown. MiR-181a is a miRNA known to regulate the threshold of TCR activation in developing thymocytes and to contribute to positive selection in the thymus [84–86]. Interestingly, miR-181a was found to be elevated upon EAE induction in lymph nodes of EAE-susceptible Dark Agouti rats as compared with the EAE-resistant rat strain Piebald Virol Glaxo [87]. Another miRNA of the same family, miR-181c, was shown to be increased in the cerebrospinal fluid of patients with MS compared with other neurological diseases, and to differentiate relapsing-remitting from secondary progressive MS courses [83]. MiR-181c‒containing extracellular vesicles were also shown to trigger the breakdown of the blood-brain barrier in a model of brain metastatic cancer [88]. Finally, miR-181b was shown to be elevated in CD4+ T cells from SLE patients, as compared with healthy controls, in a manner that was potentially linked to DNA hypomethylation in these patients [89]. To understand the biological relevance of this miRNA in regulating the activation and function of T cells, and therefore a possible role in disease onset or progression, our lab investigated the role of miR-181a in the activation of primary human T cells upon recognition of specific antigens [90]. We found that miR-181a expression regulated the threshold of activation of primary human T helper cells, especially of the Th17 type, known to be involved in MS pathogenesis [35, 37]. Moreover, we showed that, given two memory T cells bearing the same identical TCR (therefore with the same antigen recognition capacity), the cell expressing the highest levels of miR-181a became able to respond with full-blown activation and overt proliferation to the lowest concentrations of cognate antigen, regardless of the phenotype, strongly suggesting the possibility that miRNAs may also be important regulators of the threshold of T cell responses toward autoantigens in autoinflammatory diseases [90]. More studies in this direction will enable us to understand the specific roles played by these regulatory molecules in the modulation of autoimmune responses.
Interplay between DNA-modifying enzymes and miRNAs
To fully grasp the level of complexity of the mechanisms involved in fine-tuning the regulation of gene expression, one should also consider the interplay between DNA methylation and miRNA expression. Indeed, miRNAs can play a role in the modulation of DNA methylation both as targets (that is, DNA methylation can alter miRNA expression and consequently expression of downstream genes) and as effectors (that is, miRNAs reduce the expression of target DNA-modifying enzymes) (fig. 3).
For instance, the miR-29 family was shown to target DNMT1, DNMT3a and DNMT3b, leading to global DNA hypomethylation and re-expression of tumour suppressor genes in acute myeloid leukaemia cells [91]. Interestingly, miR-29 family members were also shown to be able to target TET enzymes, suggesting that this family of miRNAs acts primarily by protecting cells against perturbations in the existing DNA methylation landscape [92]. Similarly, miR-22 was shown to target directly TET proteins and to contribute to leukaemia development and metastatic potential of cancer cells [93, 94]. By utilising an unbiased high-throughput 3’ untranslated region activity screen of 460 miRNA constructs, a recent work showed that TET2 is under extensive regulation by more than 30 miRNAs (including miR-29b, miR-29c, miR-101, miR-125b, and miR-7) able to reduce TET2 expression and levels of genomic 5hmC and to disrupt normal haematopoiesis [95].
Looking at the molecular interplay between DNA methylation and miRNAs in the opposite direction, the list of miRNA genes that can be silenced by DNA methylation, especially in cancer, is constantly growing [96], and given the impact of miRNAs in regulating virtually every single aspect of T cell biology [73], it would not be surprising to identify miRNAs whose methylation-dependent silencing leads to dysregulated immune responses.
An interesting aspect of miRNA-mediated regulation is also related to the fact that selected miRNAs can be exchanged through exosomes during immune synapse formation between T cells and antigen-presenting cells (APCs) [97]. The flow of exchange was shown to be unidirectional, from the T cell to the APC, and to be able to influence gene expression in the recipient cell. Such mechanism of exchange was also shown to occur between Tregs and various immune cell types, and to suppress proliferation and cytokine secretion by pathogenic Th1 cells [98]. Importantly, miRNAs were transferred to Th1 cells both in vitroand in vivo, and such transfer significantly contributed to suppression and prevention of systemic inflammation in mice [98]. Whether exosome-mediated miRNA exchange is also able to modulate the epigenetic status of the receiving cell remains to be evaluated, and is an exciting question that will undoubtedly be addressed in work to come (box 1). Intriguingly, RNA molecules can also be methylated, and methylation of primary miRNAs (pri-miRNAs) was recently shown to act as a post-transcriptional mark promoting further miRNA processing [99]. Whether such ‘‘epitranscriptomic’’ modifications [100] will become relevant also in the context of disease will be an exciting topic for the future.
|
Box 1: Future challenges and outstanding questions‑ What is the pattern of DNA methylation and hydroxymethylation in T cells in vivo?‑ How does epigenetic variation impact disease? Is there a role for 5hmC in autoimmune diseases?‑ What are the mechanisms that dictate the activity of DNA-modifying enzymes at specific genomic loci?‑ Are there any DNA methylation-independent functions for DNMT and TET proteins?‑ Are small molecules interfering with 5hmC dynamics going to be effective and useful in the clinic?‑ Are miRNAs able to significantly alter the epigenome in vivo and in disease? |
Diet and metabolism
T cell metabolism
The role of metabolism in regulating T cell differentiation and function is becoming increasingly clear (reviewed in [101, 102]). Specifically, naïve, antigen-inexperienced T cells have a quiescent metabolism and generate energy primarily through oxidative phosphorylation (OXPHOS). Upon activation, effector T cells undergo a profound metabolic reprogramming and induce aerobic glycolysis, which is energetically less efficient, but supplies metabolic intermediates for biosynthesis of molecules required to sustain intense proliferation. Finally, memory T cells preferentially engage OXPHOS fuelled by the catabolism of fatty acids [101]. Metabolic pathways can also bias T helper cell differentiation towards specific subsets, thereby potentially influencing the outcome of an inflammatory response or autoimmune pathology [103]. Given the importance of metabolic pathways in regulating T cell responses, it is not surprising that pharmacological inhibition of some crucial metabolic regulators such as AMPK (AMP-activated protein kinase) have shown some effects in mouse models of EAE [102]. While the rodent models of EAE are extremely useful to dissect the mechanisms leading to or exacerbating disease, confirmation in the human system will be required to translate such potentially important findings to the clinic.
Dietary habits and epigenetic modifications
There is currently a great deal of interest regarding the possible link between diet, epigenetic modifications and disease. Vitamins and trace elements are important cofactors of a wide variety of enzymes as well as donors of chemical groups for epigenetic modifications. Folate is an essential B vitamin whose derivatives are main effectors in methylation reactions. Specifically, derivatives of folate are crucial to maintain appropriate methionine levels in the cells, which in turn is utilised to generate S-adenosylmethionine (SAM), the universal donor of methyl group [104]. Dietary folate intake is one of the few established links between diet and epigenetic modifications in both mouse (such as in the Agouti mouse model) and humans, as shown in rural Gambian women whose diet changes drastically according to the rainy and dry seasons [105, 106]. These studies showed that maternal folate intake is able to impact the establishment on 5mC patterns in the offspring.
Vitamin C is an essential nutrient for humans and primates, which have lost the capacity to synthesise it from glucose. Vitamin C acts as an antioxidant, able to maintain the redox state of iron-containing enzymes, and consequently to maintain the activity of dioxygenases such as TET enzymes [107]. As a result, vitamin C treatment of cultured cells led to a striking increase in the levels of genomic 5hmC, strongly influencing gene expression and differentiation potential [108]. However, whether altered levels of these cofactors can impact directly immune responses remains unclear. Vice versa, among the environmental factors known to impact the risk of MS are levels of vitamin D, whose serum levels are inversely correlated with the risk of MS. Vitamin D also appears to influence the course of MS, as lower levels are associated with increased severity of the disease [109]. Despite the fact that dietary habits and environmental factors can clearly have an influence of epigenetic modifications as well as on disease susceptibility, a precise link between these factors is still somewhat loose and the magnitude of these effects remains to be fully determined.
Epigenetic “resetting” of autoimmunity?
Although two major demethylating agents (decitabine and its analogue azacitidine) are now commonly used in the clinic for treatment of myelodysplastic syndrome and some leukaemias, the mechanisms underlying their clinical efficacy remain to be fully elucidated [110]. While genetic alterations are permanent, epigenetic modifications are potentially reversible, opening the possibility to utilise epigenetic drugs to at least relieve a certain phenotype. One of the major obstacles in this direction is, however, linked to the fact that there is still a paucity of information about the impact on disease of global changes in methylation versus gene-specific alterations. For example, despite the global hypomethylation observed in SLE patients, gene-specific hypermethylation cannot be ruled out. Moreover, the impact of other DNA modifications, such as 5hmC and its further products of oxidation, on gene transcription and disease remains for the most part to be uncovered. In general, a better understanding of the mechanisms that drive aberrant methylation and their pathological importance will be crucial to be able to exploit them for therapeutic purposes.
Disclosure statement: Work in the SM lab on this topic is supported by the Swiss National Science Foundation (31003A_156875), the San Salvatore Foundation, and the Ceresio Foundation.
References
1 Bird A. Perceptions of epigenetics. Nature. 2007;447(7143):396–8.
2 Berger SL, et al. An operational definition of epigenetics. Genes Dev. 2009;23(7):781–3.
3 Monticelli S, Natoli G. Short-term memory of danger signals and environmental stimuli in immune cells. Nat Immunol. 2013;14(8):777–84.
4 Natoli G. Maintaining cell identity through global control of genomic organization. Immunity. 2010;33(1):12–24.
5 Cullen SM, et al. Hematopoietic stem cell development: an epigenetic journey. Curr Top Dev Biol. 2014;107:39–75.
6 Okano M, et al. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999;99(3):247–57.
7 Mayle A, et al. Dnmt3a loss predisposes murine hematopoietic stem cells to malignant transformation. Blood. 2015;125(4):629–38.
8 Celik H, et al. Enforced differentiation of Dnmt3a-null bone marrow leads to failure with c-Kit mutations driving leukemic transformation. Blood. 2015;125(4):619–28.
9 Xu GL, et al. Chromosome instability and immunodeficiency syndrome caused by mutations in a DNA methyltransferase gene. Nature. 1999;402(6758):187–91.
10 Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150(1):12–27.
11 Baubec T, et al. Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature. 2015;520(7546):243–7.
12 Guo X, et al. Structural insight into autoinhibition and histone H3-induced activation of DNMT3A. Nature. 2015;517(7536):640–4.
13 Ooi SK, et al. DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature. 2007;448(7154):714–7.
14 Rishi V, et al. CpG methylation of half-CRE sequences creates C/EBPalpha binding sites that activate some tissue-specific genes. Proc Natl Acad Sci U S A. 2010;107(47):20311–6.
15 Spruijt CG, et al. Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives. Cell. 2013;152(5):1146–59.
16 Spruijt CG, Vermeulen M. DNA methylation: old dog, new tricks? Nat Struct Mol Biol. 2014;21(11):949–54.
17 Baubec T, Schubeler D. Genomic patterns and context specific interpretation of DNA methylation. Curr Opin Genet Dev. 2014;25:85–92.
18 Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet. 2012;13(7):484–92.
19 Jeltsch A. Molecular biology. Phylogeny of methylomes. Science. 2010;328(5980):837–8.
20 Heyn H, Esteller M. An Adenine Code for DNA: A Second Life for N6-Methyladenine. Cell. 2015;161(4):710–3.
21 Zemach A, et al. Genome-wide evolutionary analysis of eukaryotic DNA methylation. Science. 2010;328(5980):916–9.
22 Feng S, et al. Conservation and divergence of methylation patterning in plants and animals. Proc Natl Acad Sci U S A. 2010;107(19):8689–94.
23 Yearim A, et al. HP1 is involved in regulating the global impact of DNA methylation on alternative splicing. Cell Rep. 2015;10(7):1122–34.
24 Challen GA, et al. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet. 2012;44(1):23–31.
25 Challen GA, et al. Dnmt3a and Dnmt3b have overlapping and distinct functions in hematopoietic stem cells. Cell Stem Cell. 2014;15(3):350–64.
26 Pastor WA, Aravind L, Rao A. TETonic shift: biological roles of TET proteins in DNA demethylation and transcription. Nat Rev Mol Cell Biol. 2013;14(6):341–56.
27 Tahiliani M, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science. 2009;324(5929):930–5.
28 Ito S, et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science. 2011;333(6047):1300–3.
29 Ko M, et al. Ten-Eleven-Translocation 2 (TET2) negatively regulates homeostasis and differentiation of hematopoietic stem cells in mice. Proc Natl Acad Sci U S A. 2011;108(35):14566–71.
30 Moran-Crusio K, et al. Tet2 loss leads to increased hematopoietic stem cell self-renewal and myeloid transformation. Cancer Cell. 2011;20(1):11–24.
31 Li Z, et al. Deletion of Tet2 in mice leads to dysregulated hematopoietic stem cells and subsequent development of myeloid malignancies. Blood. 2011;118(17):4509–18.
32 Cimmino L, et al. TET family proteins and their role in stem cell differentiation and transformation. Cell Stem Cell. 2011;9(3):193–204.
33 Quivoron C, et al. TET2 inactivation results in pleiotropic hematopoietic abnormalities in mouse and is a recurrent event during human lymphomagenesis. Cancer Cell. 2011;20(1):25–38.
34 Huang Y, et al. The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing. PLoS One. 2010;5(1):e8888.
35 Simmons SB, et al. Modeling the heterogeneity of multiple sclerosis in animals. Trends Immunol. 2013;34(8):410–22.
36 Sospedra M, Martin R. Immunology of multiple sclerosis. Annu Rev Immunol. 2005;23:683–747.
37 Peters A, Lee Y, Kuchroo VK. The many faces of Th17 cells. Curr Opin Immunol. 2011;23(6):702–6.
38 Kersh EN, et al. Rapid demethylation of the IFN-gamma gene occurs in memory but not naive CD8 T cells. J Immunol. 2006;176(7):4083–93.
39 Makar KW, et al. Active recruitment of DNA methyltransferases regulates interleukin 4 in thymocytes and T cells. Nat Immunol. 2003;4(12):1183–90.
40 Bruniquel D, Schwartz RH. Selective, stable demethylation of the interleukin-2 gene enhances transcription by an active process. Nat Immunol. 2003;4(3):235–40.
41 Gamper CJ, et al. Identification of DNA methyltransferase 3a as a T cell receptor-induced regulator of Th1 and Th2 differentiation. J Immunol. 2009;183(4):2267–76.
42 Lee PP, et al. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity. 2001;15(5):763–74.
43 Yu Q, et al. DNA methyltransferase 3a limits the expression of interleukin-13 in T helper 2 cells and allergic airway inflammation. Proc Natl Acad Sci U S A. 2012;109(2):541–6.
44 Hutchins AS, et al. Gene silencing quantitatively controls the function of a developmental trans-activator. Mol Cell. 2002;10(1):81–91.
45 Makar KW, Wilson CB. DNA methylation is a nonredundant repressor of the Th2 effector program. J Immunol. 2004;173(7):4402–6.
46 Scharer CD, et al. Global DNA methylation remodeling accompanies CD8 T cell effector function. J Immunol. 2013;191(6):3419–29.
47 Hashimoto S, et al. Coordinated changes in DNA methylation in antigen-specific memory CD4 T cells. J Immunol. 2013;190(8):4076–91.
48 Ichiyama K, et al. The Methylcytosine Dioxygenase Tet2 Promotes DNA Demethylation and Activation of Cytokine Gene Expression in T Cells. Immunity. 2015.
49 Lal G, Bromberg JS. Epigenetic mechanisms of regulation of Foxp3 expression. Blood. 2009;114(18):3727–35.
50 Feng Y, et al. Control of the inheritance of regulatory T cell identity by a cis element in the Foxp3 locus. Cell. 2014 ;158(4):749–63.
51 Polansky JK, et al. DNA methylation controls Foxp3 gene expression. Eur J Immunol. 2008;38(6):1654–63.
52 Wang L, et al. Foxp3+ T-regulatory cells require DNA methyltransferase 1 expression to prevent development of lethal autoimmunity. Blood. 2013;121(18):3631–9.
53 International Multiple Sclerosis Genetics, C., et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature. 2011;476(7359):214–9.
54 Willer CJ, et al. Twin concordance and sibling recurrence rates in multiple sclerosis. Proc Natl Acad Sci U S A. 2003;100(22):12877–82.
55 Ballestar E, Esteller M, Richardson BC. The epigenetic face of systemic lupus erythematosus. J Immunol. 2006;176(12):7143–7.
56 Brodin P, et al. Variation in the human immune system is largely driven by non-heritable influences. Cell. 2015;160(1-2):37–47.
57 Baranzini SE, et al. Genome, epigenome and RNA sequences of monozygotic twins discordant for multiple sclerosis. Nature. 2010;464(7293):1351–6.
58 Farh KK, et al. Genetic and epigenetic fine mapping of causal autoimmune disease variants. Nature. 2015;518(7539):337–43.
59 Rakyan VK, et al. Identification of type 1 diabetes-associated DNA methylation variable positions that precede disease diagnosis. PLoS Genet. 2011;7(9): p. e1002300.
60 Quddus J, et al. Treating activated CD4+ T cells with either of two distinct DNA methyltransferase inhibitors, 5-azacytidine or procainamide, is sufficient to cause a lupus-like disease in syngeneic mice. J Clin Invest. 1993;92(1):38–53.
61 Hedrich CM, et al. Stat3 promotes IL-10 expression in lupus T cells through trans-activation and chromatin remodeling. Proc Natl Acad Sci U S A. 2014;111(37):13457–62.
62 Rauen T, et al. cAMP-responsive element modulator (CREM)alpha protein induces interleukin 17A expression and mediates epigenetic alterations at the interleukin-17A gene locus in patients with systemic lupus erythematosus. J Biol Chem. 2011;286(50):43437–46.
63 Picascia A, et al. Epigenetic control of autoimmune diseases: from bench to bedside. Clin Immunol. 2015;157(1):1–15.
64 Glant TT, Mikecz K, Rauch TA. Epigenetics in the pathogenesis of rheumatoid arthritis. BMC Med. 2014;12:35.
65 Esteller M. Epigenetics in cancer. N Engl J Med. 2008;358(11):1148–59.
66 Lian CG, et al. Loss of 5-hydroxymethylcytosine is an epigenetic hallmark of melanoma. Cell. 2012;150(6):1135–46.
67 Leoni C, et al. Reduced DNA methylation and hydroxymethylation in patients with systemic mastocytosis. Eur J Haematol. 2015.
68 Ko M, et al. Impaired hydroxylation of 5-methylcytosine in myeloid cancers with mutant TET2. Nature. 2010;468(7325):839–43.
69 Smith ZD, Meissner A. DNA methylation: roles in mammalian development. Nat Rev Genet. 2013;14(3):204–20.
70 Issa JP. Aging and epigenetic drift: a vicious cycle. J Clin Invest. 2014;124(1):24–9.
71 Maegawa S, et al. Widespread and tissue specific age-related DNA methylation changes in mice. Genome Res. 2010;20(3):332–40.
72 Niwa T, et al. Inflammatory processes triggered by Helicobacter pylori infection cause aberrant DNA methylation in gastric epithelial cells. Cancer Res. 2010;70(4):1430–40.
73 Monticelli S. MicroRNAs in T helper cell differentiation and plasticity. Semin Immunol. 2013;25(4):291–8.
74 Pauley KM, Cha S, Chan EK. MicroRNA in autoimmunity and autoimmune diseases. J Autoimmun. 2009;32(3-4):189–94.
75 Wei B, Pei G. microRNAs: critical regulators in Th17 cells and players in diseases. Cell Mol Immunol. 2010;7(3):175–81.
76 Pan W, et al. MicroRNA-21 and microRNA-148a contribute to DNA hypomethylation in lupus CD4+ T cells by directly and indirectly targeting DNA methyltransferase 1. J Immunol. 2010;184(12):6773–81.
77 Murugaiyan G, et al. Silencing microRNA-155 ameliorates experimental autoimmune encephalomyelitis. J Immunol. 2011;187(5):2213–21.
78 Smith KM, et al. miR-29ab1 deficiency identifies a negative feedback loop controlling Th1 bias that is dysregulated in multiple sclerosis. J Immunol. 2012;189(4):1567–76.
79 Du C, et al. MicroRNA miR-326 regulates TH-17 differentiation and is associated with the pathogenesis of multiple sclerosis. Nat Immunol. 2009;10(12):1252–9.
80 Zhu E, et al. miR-20b suppresses Th17 differentiation and the pathogenesis of experimental autoimmune encephalomyelitis by targeting RORgammat and STAT3. J Immunol. 2014;192(12):5599–609.
81 Lindberg RL, et al. Altered expression of miR-17-5p in CD4+ lymphocytes of relapsing-remitting multiple sclerosis patients. Eur J Immunol. 2010;40(3):888–98.
82 Reijerkerk A, et al. MicroRNAs regulate human brain endothelial cell-barrier function in inflammation: implications for multiple sclerosis. J Neurosci. 2013;33(16):6857–63.
83 Haghikia A, et al, Regulated microRNAs in the CSF of patients with multiple sclerosis: a case-control study. Neurology. 2012;79(22):2166–70.
84 Ebert PJ, et al. An endogenous positively selecting peptide enhances mature T cell responses and becomes an autoantigen in the absence of microRNA miR-181a. Nat Immunol. 2009;10(11):1162–9.
85 Li QJ, et al. miR-181a is an intrinsic modulator of T cell sensitivity and selection. Cell. 2007;129(1):147–61.
86 Neilson JR, et al. Dynamic regulation of miRNA expression in ordered stages of cellular development. Genes Dev. 2007;21(5):578–89.
87 Bergman P, et al. Next-generation sequencing identifies microRNAs that associate with pathogenic autoimmune neuroinflammation in rats. J Immunol. 2013;190(8):4066–75.
88 Tominaga N, et al. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood-brain barrier. Nat Commun. 2015;6:6716.
89 Zhao M, et al. DNA methylation and mRNA and microRNA expression of SLE CD4+ T cells correlate with disease phenotype. J Autoimmun. 2014;54:127–36.
90 Mele F, et al. ERK phosphorylation and miR-181a expression modulate activation of human memory T17 cells. Nat Commun. 2015;6:6431.
91 Garzon R, et al. MicroRNA-29b induces global DNA hypomethylation and tumor suppressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood. 2009;113(25):6411–8.
92 Morita S, et al. miR-29 represses the activities of DNA methyltransferases and DNA demethylases. Int J Mol Sci. 2013;14(7):14647–58.
93 Song SJ, et al. MicroRNA-antagonism regulates breast cancer stemness and metastasis via TET-family-dependent chromatin remodeling. Cell. 2013;154(2):311–24.
94 Song SJ, et al. The oncogenic microRNA miR-22 targets the TET2 tumor suppressor to promote hematopoietic stem cell self-renewal and transformation. Cell Stem Cell. 2013;13(1):87–101.
95 Cheng J, et al. An extensive network of TET2-targeting MicroRNAs regulates malignant hematopoiesis. Cell Rep. 2013;5(2):471–81.
96 Lopez-Serra P, Esteller M. DNA methylation-associated silencing of tumor-suppressor microRNAs in cancer. Oncogene. 2012;31(13):1609–22.
97 Mittelbrunn M, et al. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat Commun. 2011;2:282.
98 Okoye IS, et al. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity. 2014;41(1):89–103.
99 Alarcon CR, et al. N6-methyladenosine marks primary microRNAs for processing. Nature. 2015;519(7544):482–5.
100 Lee M, Kim B, Kim VN. Emerging roles of RNA modification: m(6)A and U-tail. Cell. 2014;158(5):980–7.
101 O’Sullivan D, Pearce EL. Targeting T cell metabolism for therapy. Trends Immunol. 2015;36(2):71–80.
102 Lochner M, Berod L, Sparwasser T. Fatty acid metabolism in the regulation of T cell function. Trends Immunol. 2015;36(2):81–91.
103 Berod L, et al. De novo fatty acid synthesis controls the fate between regulatory T and T helper 17 cells. Nat Med. 2014;20(11):1327–33.
104 Moen EL, et al. New themes in the biological functions of 5-methylcytosine and 5-hydroxymethylcytosine. Immunol Rev. 2015;263(1):36–49.
105 Dominguez-Salas P, et al. Maternal nutrition at conception modulates DNA methylation of human metastable epialleles. Nat Commun. 2014;5:3746.
106 Jirtle RL, Skinner MK. Environmental epigenomics and disease susceptibility. Nat Rev Genet. 2007;8(4):253–62.
107 Monfort A, Wutz A. Breathing-in epigenetic change with vitamin C. EMBO Rep. 2013;14(4):337–46.
108 Blaschke K, et al. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature. 2013;500(7461):222–6.
109 Koch MW, Metz LM, Kovalchuk O. Epigenetics and miRNAs in the diagnosis and treatment of multiple sclerosis. Trends Mol Med. 2013;19(1):23–30.
110 Christman JK. 5-Azacytidine and 5-aza-2'-deoxycytidine as inhibitors of DNA methylation: mechanistic studies and their implications for cancer therapy. Oncogene. 2002;21(35):5483–95.